To access and download PowerPoint presentation on 'Regulation of Floral Development' click on the link below:
PHYTOCHROME AND PHOTOPERIODISM
Phytochrome


Phytochrome is a dimer composed of two polypeptides
•Native phytochrome occurs as a homo-dimer.
•Each subunit consists of:
•a light-absorbing pigment molecule called the chromophore
•a polypeptide chain called the apoprotein.
•The chromophore is attached to the apoprotein through a thioether linkage to a cysteine residue
•Both the chromophore and protein undergo conformational changes during Pr-Pfr interconversion.
PHY genes encode two types of phytochrome:
•Phytochrome is encoded by a multigene family consisting of five members: PHYA, PHYB, PHYC, PHYD, and PHYE.
Type I phytochrome:
•Encoded by PHYA gene
•Transcriptionally active in dark-grown seedlings
•Expression is strongly inhibited in light in monocots and less dramatically in dicots.
•PfrA is also unstable.

Type II phytochrome:
•Encoded by PHYB, PHYC, PHYD, PHYE genes
•Expression of their mRNAs is not significantly changed by light.
•Proteins are more stable in the Pfr form.

Phytochrome is the primary photoreceptor in photoperiodism
Reason:

A Blue-Light Photoreceptor Also Regulates Flowering
Five genetically distinct developmental pathways control flowering. These are:
Photoperiodic pathway
It involves phytochromes (PHYA and PHYB acting antagonistically) and cryptochromes.
In Arabidopsis (LDP)
•In the morning, PHYB represses CONSTANS (CO) gene expression.
•In the evening, PHYA and cryptochrome interacts with circadian clock genes to induce CO expression.
•CO is a transcription factor which induces FLOWERING LOCUS T (FT).
•FT protein (‘Florigen’) travels from the induced leaf to the shoot apex via the phloem.
•FT forms a complex with transcription factor FD.
•FT-FD complex activates downstream genes like SOC1, LFY, etc which in turn activates the floral homeotic genes for floral development.
In rice (SDP)
•In the morning, PHYB represses Heading date 1 (Hd1) which is the homologue of CO.
•In the evening, PHYA interacts with circadian clock genes to induce Hd1 expression.
•Here, Hd1 acts as an inhibitor of flowering.
•Absence of Hd1 stimulates expression of Hd3a.
•Hd3a (‘Florigen’) travels from the induced leaf to the shoot apex via phloem for floral development.

Coincidence model
•Transition to flowering occurs when the expression peak of CO coincides with the light phase.
•PHYA/PHYB perceive light signals and entrain the clock
•The clock genes control the diurnal expression of CO and its expression peaks between 12 hours of light and dawn (SuaÂrez-LoÂpez et al., 2001; Nature).
•In long days, the CO expression peak coincides with the light phase (coincidence phase).
•In light, the CO protein is stable and can activate FT expression, promoting flowering.
•In short days, CO expression peaks in the dark (no coincidence of light phase and CO expression).
•In the dark, CO protein is degraded and thus cannot induce FT expression and further processes.

(Singh et al., 2016; New Phytologists)
Florigen concept
•During flowering, a biochemical signal is produced in photoperiodically induced leaves and is then transmitted to the shoot apex where it stimulates flowering.
•This mode of action resembles a hormonal effect.
•Hence, Mikhail Chailakhyan postulated the existence of a universal flowering hormone, florigen (1930s).
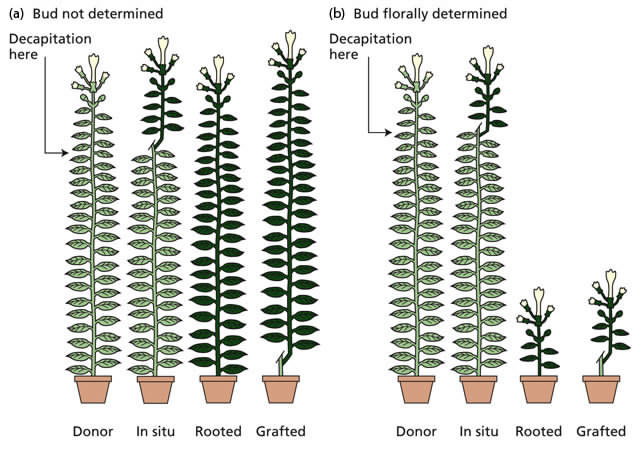
•Evidence: non-induced receptor plants were stimulated to flower by being grafted onto a leaf or shoot from photoperiodically induced donor plants.
•Movement of the floral stimulus from a donor leaf to the stock across the graft union correlated closely with the translocation of 14C-labeled assimilates via vascular continuity across the graft union.
Identification of florigen
•Since 1930s, many attempts to isolate florigen have remained unsuccessful.
•A major breakthrough is the identification of FLOWERING LOCUS T (FT) in Arabidopsis.
•FT is a downstream target of CO and is expressed in companion cells of leaves (like CO).
•FT is a small globular protein induced by CO in response to long days.
•FT then travels from leaf to shoot apex via phloem to induce flowering.
•It can travel through grafting unions and induce flowering in competent recipient plants.
•Therefore, FT (or its homologue, Hd3a in rice) is the florigen.
Regulation of floral induction by FT
•FT mRNA is synthesized in leaf companion cells.
•Endoplasmic reticulum protein, FT INTERACTING PROTEIN 1 (FTIP1), helps the transport of FT from companion cells to sieve tubes.
•FT protein then moves via phloem translocation stream to shoot apex.
•In the shoot apex (floral meristem), FT enters the nucleus to bind FLOWERING LOCUS D (FD) which is a transcription factor.
•FT-FD complex activates floral identity genes like SOC1 and AP1.
•SOC1 then activates LFY which further activates the floral homeotic genes for floral development.




Sucrose pathway
•Sucrose stimulates flowering in Arabidopsis by increasing LFY expression.
Gibberellin pathway
•Required for early flowering and for flowering under non-inductive short days.
•GA induces GA-MYB (transcription factor) mediated activation of LFY.

(Images kindly shared by Oxford University Press and are being used for teaching purpose only)